
6: Biofuels B - Cellulosic Ethanol
- Page ID
- 34461
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)(Learning goals written by Claude, Anthropic)
By the end of this chapter, students should be able to:
Cellulose Structure and Stability
- Explain the three reinforcing reasons cellulose resists enzymatic hydrolysis — the intrinsic stability of the β(1,4) glycosidic bond (t₁/₂ ≈ 5 million years uncatalyzed), crystalline packing of cellulose Iα and Iβ polymorphs stabilized by intralayer hydrogen bonds and interlayer hydrophobic planes from axial C–H atoms, and encasement by lignin in the secondary cell wall — and contrast this with the much greater reactivity of starch's α(1,4) bonds.
- Describe the composition and organization of the primary and secondary plant cell walls — including the roles of cellulose microfibrils (40–60%), hemicellulose (20–40%), pectin, and lignin — and explain why most bioethanol-relevant cellulose comes from the secondary cell wall of woody plants and switchgrass.
Lignin Structure and Degradation
- Describe how the three monolignols (p-coumaryl, coniferyl, sinapyl alcohols) are derived from phenylalanine via cinnamic acid, and explain how peroxidase-catalyzed phenoxide radical formation drives oxidative coupling to produce the heterogeneous, covalently cross-linked lignin polymer.
- Describe the four-copper active site of fungal laccase from Trametes versicolor (T1, T2, T3α, T3β), explain how four sequential single-electron transfers oxidize lignin aromatic substrates with O₂ as the terminal electron acceptor producing water, and identify the key substrate-binding residues (His-458, Asp-206, Asn-264, Phe-265) and why thermochemical pretreatment is required before enzymatic delignification can be effective.
Cellulase Biochemistry and the Processive Cycle
- Distinguish the inversion mechanism (single SN2-like water attack, two carboxylate general acid/base catalysts) from the retention mechanism (double-displacement via a glycosyl-enzyme intermediate) of glycoside hydrolases, predict the stereochemical outcome at the anomeric carbon for each, and identify which mechanism TrCel7A uses with Glu217 as nucleophile and Glu212/Asp214 as acid/base catalysts.
- Describe the modular architecture of TrCel7A — catalytic domain (CD) with a 50 Å substrate tunnel, glycosylated linker, and C-terminal carbohydrate-binding module (CBM) — and explain how each component contributes to the processive cycle of cellulose chain threading, hydrolysis, cellobiose expulsion, and eventual desorption.
- Explain how the triplet tyrosines (Y5, Y31, Y32) of the CBM bind to the hydrophobic face of crystalline cellulose through π-stacking and van der Waals interactions, how N- and O-glycosylation of the linker enhances cellulose binding, and why nonproductive adsorption of the same CBM surface to lignin reduces effective cellulase concentration available for catalysis.
Introduction
In the last section, we explored how ethanol can be made from corn starch, an α(1,4) polymer of glucose with α(1,6) branches. Its production, however, comes at a cost. Recent life-cycle studies have shown that, compared to fossil fuels, corn ethanol releases as much, if not more, CO2. In addition, is it ethically justifiable to remove so much land from food production to produce bioethanol that, at present, is worse than fossil fuels from a climatic perspective?
To address these issues, significant work has been done to produce ethanol from cellulose, a β(1,4) polymer of glucose, the most abundant biomolecule on Earth. Cellulose from trees, switchgrass, and "waste biomass" are prime sources for bioethanol production. Waste biomass includes stover (field crop stalks and leaves), straw, wood chips, and sawdust. About 113 gallons of ethanol can be made from one ton of corn stover, close to the 124 gallons produced from corn.
Nature routinely breaks down cellulose using cellulases, enzymes found in bacteria, fungi, protozoans, plants, and some animals. Ruminants and even termites obtain cellulases from microbes living within their guts. The fungal-mediated decay of dead trees requires microbial cellulase,s but think how slow that process is. This stems partly from the very strong β(1,4) glycosidic link connecting glucose monomers in the polymer, which, in the absence of a catalyst and at neutral pH, has an estimated half-life of 5 million years. Fossilized plants have been found to have intact cellulose and chitin, a β(1,4) polymer of N-acetylglucosamine. The β(1,4) glycosidic link in cellulose is orders of magnitude more stable than the phosphodiester bond of nucleic acids and the amide link of proteins. They are, however, readily cleaved by glycosidases such as cellulases, which can increase the kcat/KM over the uncatalyzed rate by up to 1017 fold, even in the absence of active site metal ions to facilitate hydrolysis (reference).
Another reason for the slow decay of dead trees is the complex structure of the cell wall, particularly the presence of another tree polymer, lignin. Lignin stabilizes the cell wall and creates substantial barriers to cellulose's access to additional glycosidases.
A final reason for cellulose's extreme stability is the "quaternary" structure of the β(1,4)-linked cellulose strands, which consist of densely packed, intertwined strands that limit solvent (in this case, water) accessibility necessary for hydrolysis. In addition, some exposed surface planes of the packed cellulose strands are hydrophobic. This might seem startling, given the polymer's polar nature. Let's review it now since our goal is to present climate change from a biochemical perspective! Some of this material has been presented in previous chapters, but we will reuse it here so this chapter section can stand alone.
A review: The Plant Cell Wall
(See Chapter 7.3 for more details.) There are about 35 types of plant cells, and each may have a different cell wall depending on the local needs of that cell. Cells synthesize a thin cell wall that extends and stays thin as the cell grows. Figure \(\PageIndex{1}\) shows the primary cell wall of plants. The primary cell wall contains cellulose microfibrils (no surprise) and two other polymers, pectin and hemicellulose. The middle lamella, which consists of pectins, is somewhat analogous to the extracellular matrix.
 Figure \(\PageIndex{1}\): Primary cell wall of plants. https://commons.wikimedia.org/wiki/F...diagram-en.svg
Figure \(\PageIndex{1}\): Primary cell wall of plants. https://commons.wikimedia.org/wiki/F...diagram-en.svg
After cell growth, the cell often synthesizes a secondary cell wall, thicker than the primary wall, for greater rigidity. Since the enzymatic machinery for its synthesis is in the cytoplasm and the cell membrane, it is deposited between the cell membrane and the primary cell wall, as shown in the animated image in Figure \(\PageIndex{2}\).
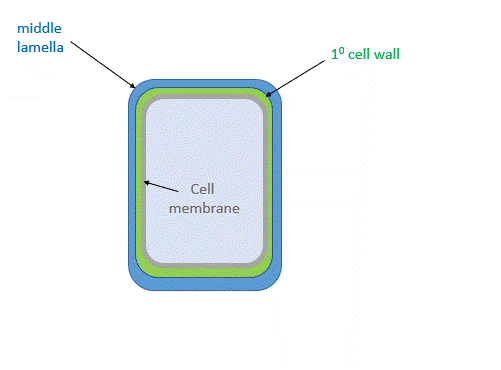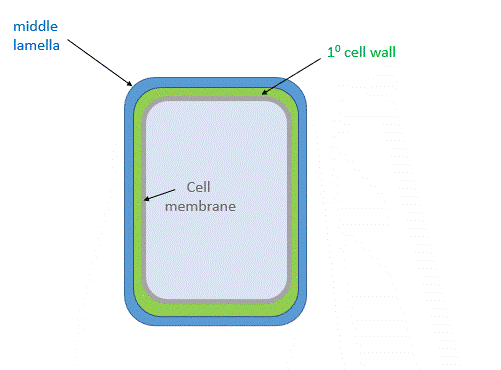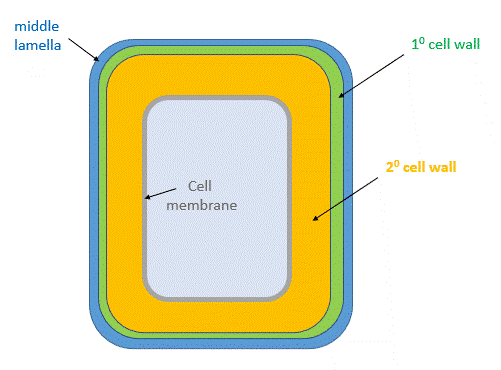 Figure \(\PageIndex{2}\): Primary and secondary cell wall of plants
Figure \(\PageIndex{2}\): Primary and secondary cell wall of plants
Figure \(\PageIndex{3}\) shows a structural representation of the primary and secondary cell walls.
 Figure \(\PageIndex{3}\): structural representation of both the primary and secondary cell walls of plants. Nakano Yoshimi et al. Frontiers in Plant Science (6), 288 (2015) https://www.frontiersin.org/article/...015.00288/full. Creative Commons AttributionLicense (CC BY).
Figure \(\PageIndex{3}\): structural representation of both the primary and secondary cell walls of plants. Nakano Yoshimi et al. Frontiers in Plant Science (6), 288 (2015) https://www.frontiersin.org/article/...015.00288/full. Creative Commons AttributionLicense (CC BY).
The middle lamella, which contains pectins, lignins, and some proteins, helps "glue together" the primary cell walls of surrounding plants.
Primary Cell Wall:
The main component of the primary plant wall is the homopolymer cellulose (40% -60% mass) in which the glucose monomers are linked β(1→4)-linked into strands that collect into microfibrils through hydrogen bond interactions. Two other groups of polymers, hemicellulose and pectin, make up the plant cell wall.
Hemicellulose can make up 20-40% of the mass. These polymers have β(1,4) backbones of glucose, mannose, or xylose (called xyloglucans, xylans, mannans, galactomannans, glucomannans, and galactoglucomannans along with some β(1,3 and 1,4)-glucans. The most abundant hemicellulose in higher plants is the xyloglucans, which have a cellulose backbone linked at O6 to α-D-xylose. Pectin consists of linked galacturonic acids forming homogalacturonans, rhamnogalacturonans, and rhamnogalacturonans II (RGII) [12] [13]. Homogalacturonans (α1→4) linked D-GalA make up more than 50% of the pectin. Figure \(\PageIndex{4}\) shows some structures. They are generally branched, shorter than cellulose chains, and can often crystallize.

Figure \(\PageIndex{4}\): Variant of the cell wall components of a plant. Costa and Plazanet. Advances in Biological Chemistry 06(03):70-105. DOI: 10.4236/abc.2016.63008. License CC BY 4.0
Secondary Cell Wall
The structure of the secondary cell wall depends on the cell's function and environment. It contains cellulose fibers, hemicellulose, and a new polymer, lignin. It is abundant in xylem vessels and fiber cells of woody plants. It gives the plant extra stability and new functions, including the transport of fluids through channels. The proportion of cellulose in the secondary cell wall is higher than in the primary cell wall, and it is less hydrated than in the primary cell wall. Given the relative volume of the secondary and primary cell walls inferred from Fig. 2, most of the tree-derived cellulose for bioethanol production comes from the secondary cell wall. Switch grasses, a perennial plant, are also valuable sources of cellulose (32–45% wt) and hemicellulose (21–31% wt), but they also contain significant amounts of lignin (12–28% wt). In summary, the secondary cell wall, formed after the cell stops growing, accounts for most of the carbohydrate biomass of plants.
Glycosidases, mostly α- and β-amylases, are needed to convert corn-derived starch into glucose for fermentation and ethanol production. Likewise, cellulases are required to degrade cellulose into glucose for the production of cellulosic ethanol. However, it is a much more complex process since most cellulose is in the secondary cell wall. The lignin barrier in the walls protects cellulose from accessibility to cellulases, even after chemical and thermal pre-processing. In addition, xylans, which can make up 30% of the mass of the secondary cell wall, also inhibit cellulose degradation.
A thermochemical process can convert cellulose to the synthetic gases CO and H2, which can be used as reactants to form ethanol. We'll discuss the biochemical process that uses pretreatment and enzymatic hydrolysis to produce cellulosic ethanol. Lignin can be recovered and used to provide energy for the industrial-scale synthesis of cellulosic ethanol.
Let's explore the barriers lignin poses and how they can be surmounted to facilitate access to cellulose and the liberation of glucose for cellulosic ethanol production.
Lignin Structure and Reactivity
Lignins, which can make up to 25% of the biomass weight of secondary walls, are made from phenylalanine derivatives but more directly from cinnamic acid. This is made from hydroxylated phenylalanine and converted through other steps to hydroxycinnamic alcohols called monolignols, as shown in Figure \(\PageIndex{5}\). Three typical monomers, p-coumaryl, coniferyl, and sinapyl alcohols, can polymerize into lignins, with their units in the polymer (P) named hydroxyphenyl, guaiacyl, and syringly, respectively.

Lignols are activated phenolic compounds that form phenoxide free radicals (catalyzed by peroxidase enzymes), which can attack a second lignol to form covalent dimers. Reaction mechanisms for dimerizing the MS sinapyl alcohol-free radical are shown in Figure \(\PageIndex{6}\).

Figure \(\PageIndex{6}\): Dimers of lignols
Now, imagine this polymerization continuing, forming more phenolic free radicals and coupling at multiple sites to form a large covalent lignin polymer. Figure \(\PageIndex{7}\) shows one example of a larger lignin.

Figure \(\PageIndex{7}\): A larger lignin. https://commons.wikimedia.org/wiki/C...ile:Lignin.png . By Smokefoot - Own work, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=16022799
Lignin strengthens the cell wall and further stabilizes the already unreactive cellulose fibers. Let's look at a specific example - using corn stover (CS) as a cellulose source. This process includes chemical pretreatment of the biomass source and the addition of a bacterial strain, Pandoraea sp. B-6 (B-6) isolated from long, narrow strips of bamboo (slips). Bamboo is a type of woody grass that grows rapidly. These bacteria produce two extracellular lignin-degrading enzymes, manganese peroxidase (MnP) and laccase (Lac). Laccase (Lac) is a multi-copper oxidase that uses O2 to oxidize the syringyl, guaiacyl, and p-hydroxyphenyl monomers in lignins. MnP has similar properties. These and other enzymes can lead to the depolymerization of lignin and the degradation of lignin-derived aromatic compounds
The addition of the B-6 bacteria (a source of MnP and Lac) to milled corn stover (CS) did not increase the rate of lignin degradation unless the corn stover was preincubated with a tetrahydrofuran-water (THF–H2O) solution with 0.5 wt% sulfuric acid and heated to 150 oC. This led to the erosion of the corn stover, allowing access to the bacterial enzyme. The untreated and pre-treated CS surface, along with a diagram showing access of Lac and MnP to the lignin, is shown in Figure \(\PageIndex{8}\).

Figure \(\PageIndex{8}\): Untreated and pre-treated CS surface and Lac and MnP interaction with lignins. Zhuo, S., Yan, X., Liu, D. et al. Use of bacteria for improving the lignocellulose biorefinery process: the importance of pre-erosion. Biotechnol Biofuels 11, 146 (2018). https://doi.org/10.1186/s13068-018-1...068-018-1146-4. Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/)
In addition to restricting cellulase's access to cellulose, it can also nonspecifically adsorb to lignin and its pretreated forms. The lignin derivatives exhibit a more hydrophobic surface, which promotes cellulase interactions. Some plant laccases are involved in lignin biosynthesis, but in bacteria and fungi, they are likely involved in degradation.
Of course, fungi, which are prime degraders of dead biomass in forests, are also sources of enzymes for lignin degradation. For example, species of white rot fungi produce manganese peroxidase (MnP), lignin peroxidase (LiP), versatile peroxidase (VP), and laccase (Lac). They form reactive lignin-derived aromatic free radicals (similar to those produced in lignin synthesis), leading to the breaking of ether bonds, aromatic ring cleavage, and the removal of methoxy groups from the substrate, a process called delignification. Pretreatment of the biomass increases the yields of available cellulose. Fungi, however, grow slowly, and the delignification rate is still low. In addition, they have hydrolytic enzymes that reduce cellulose yield. That is why bacterial sources like B6 are sought for delignification.
As this is a biochemistry textbook, let's explore the structure and function of fungal laccase. The enzyme can bind many hydroxylated and methoxy-aromatic compounds as substrates, so its active site must be adaptable and likely dynamic. The laccase (TvL) from the fungus Trametes versicolor has been used for structural analyses, in-silico docking experiments, and molecular dynamics simulations.
The enzyme contains four copper ions at a T1 Cu site and a trinuclear Cu cluster (T2 Cu, T3α Cu, and T3β Cu) at a T2/T3 site. As the mechanism involves free radical intermediates with O2 as both oxidant and substrate, 4 electrons are transferred in single-electron steps to the T1 Cu, then to the other three coppers, and finally to O2, forming 2 water molecules as products. The amino acid side chain ligands ligating four copper ions are shown for white rot fungi laccase from Trametes Versicolor in Figure \(\PageIndex{9}\).

Figure \(\PageIndex{9}\): T1 Cu (top left) and the trinuclear Cu cluster (T2, T3α and T3β) and their ligands for Trametes Versicolor laccase (TvL, pdb: 1GYC)
Fungal laccases are extracellular proteins with about 550 amino acids arranged in three cupredoxin-like, beta-barrel domains. The T1 Cu is close to the surface and is found in domain 3, while the other copper ions are buried at the interface between domains 1 and 3. Figure \(\PageIndex{10}\) shows an interactive iCn3D modelof Laccase from the Fungus Trametes Versicolor (1GYC)
.png?revision=1&size=bestfit&width=430&height=434)
 Figure \(\PageIndex{10}\): Laccase from the Fungus Trametes Versicolor (1GYC). (Copyright; author via source).
Figure \(\PageIndex{10}\): Laccase from the Fungus Trametes Versicolor (1GYC). (Copyright; author via source).
Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...cSYir86P9nxSW6
Domain 1 is green, domain 2 magenta, and domain 3, which contains the single T1 Cu, is orange. The protein is glycosylated, as shown in the blue glycan cube cartoons representing N-acetylglucosamine. Key substrate binding and catalytic side chains are shown in sticks and labeled; Asp 206 is a critical residue involved in substrate binding.
The binding interactions of TvL with various aromatic substrates are shown schematically in Figure \(\PageIndex{11}\).

Figure \(\PageIndex{11}\): Binding modes of representative compounds for TvL. Mehra, R., Muschiol, J., Meyer, A.S. et al. A structural-chemical explanation of fungal laccase activity. Sci Rep 8, 17285 (2018). https://doi.org/10.1038/s41598-018-35633-8. Creative Commons Attribution 4.0 International License. http://creativecommons.org/licenses/by/4.0/
Most substrates interact with the highly conserved His-458 (blue color, ligand for the Ti Cu)and Asp-206 (orange color) residues and form hydrogen bonds, salt bridges, or π-π stacking interactions. Asn-264 (blue) and Phe-265 (green) form important hydrogen bonding and π-π stacking interactions with substrates. The green color highlights nonpolar side chains. Ligands bind near the Ti Cu in domain 3 to initiate electron transfer. The active site of TvL must be dynamic to bind the various-sized ligands shown above. Molecular dynamics simulations, shown in Figure \(\PageIndex{12}\), support this.

Figure \(\PageIndex{12}\): Display of molecular dynamics simulations showing the loop regions of TvL (magenta colored) and another laccase, CuL (yellow colored), and high levels of fluctuations. Mehra, R., ibid.
Breaking down Cellulose
We just explained how the lignin barrier could be degraded to allow cellulase to access cellulose. As we described above, this also poses a difficult challenge given the stability and inaccessibility of the glucosidic bonds in cellulose. The inaccessibility of "naked" cellulose fibers stems partly from the tight binding of cellulose strands into crystal lattices. Multiple crystal forms of cellulose, called polymorphs, can form. Plant cellulose has two predominant polymorphs, cellulose Iβ and Iα. Their structures are shown below in Figure \(\PageIndex{13}\).

Figure \(\PageIndex{13}\): Natural and synthetic cellulose polymorphs. Christina M. Payne et al. Chem. Rev. 2015, 115, 3, 1308–1448 (2015), https://doi.org/10.1021/cr500351c. Open access through a Creative Commons public use license.
They both form hydrogen bonds within a layer, with the main differences resulting from interlayer stacking. There are no hydrogen bonds between layers. You might find that surprising at first glance, but remember that all the OH groups in the lowest-energy chair form of glucose are equatorial, which allows intralayer hydrogen bonding. The interactions between layers predominantly arise from van der Waals interactions, specifically induced dipole-induced dipole interactions. The hydrophobic planes, arising from axial H atoms projecting above and above each planar layer of the cellulose fibers, can be readily seen in Figure \(\PageIndex{14}\).

Figure \(\PageIndex{14}\): Hydrophobic planes arising from axial H atoms projecting above and above each planar layer of the cellulose fibers. Akira Isogai et al. Progress in Polymer Science, 86 (2018), https://doi.org/10.1016/j.progpolymsci.2018.07.007. Creative Commons license
Now, we can explore the structure of cellulases and how they bind to and cleave cellulose.
Cellulases, which cleave β(1,4) glycosidic bonds in cellulose, are members of a family of enzymes that go by many names, including glycosidases or, more recently, glycoside hydrolases (GH). The Carbohydrate Active Enzymes (CAZypedia) has over 128 glycoside hydrolase (GH) family web pages, with enzymes that form hemiacetals during glycosidic bond cleavage. Fungal cellulases that act on cellulose are found in GH families 5, 6, 7, 12, and 45.
There are many types of secreted or cell-surface cellulases, including endoglucanases, exoglucanases (example is cellobiohydrolases (CBHs), and β-glucosidase (BG) ). We will focus on cellobiohydrolases (CBHs), the most studied one, which cleaves a 2-glucose unit (cellobiose) from either end of cellulose as it proceeds (processes) along the chain. Fungal and bacterial CBHs can also act on crystalline cellulose. The resulting cellobiose is further cleaved by β-glucosidases. Ruminants and even termites obtain cellulases from microbes living within their guts. The enzyme has a "tunnel" between two surface loops, which interacts with and processively cleaves cellulose.
As mentioned above, fungi are the major degraders of biomass and are critical to the carbon cycle. Some fungi(brown-rot) use the Fenton reaction (Chapter 13.3) to produce the very reactive hydroxyl free radical (.OH), which causes biomass degradation. Filamentous fungi (such as white and soft rot and T. reesei) use enzymes. The T. reesei cellulase, cellobiose hydrolase 1, is now called TrCel7A because it belongs to the GH 7A family.
Cellulase mechanism.
We have previously described the mechanisms of polysaccharide synthesis (Chapter 20.3), so we will discuss the mechanism of the very similar reaction of cellulose degradation by cellulase in less detail. Two general mechanisms are possible: one leading to retention of configuration at the resulting hemiacetal end of cellobiose, and another leading to inversion of configuration. These mechanisms are shown in Figure \(\PageIndex{15}\) for glucose in alpha-linkage at the anomeric carbon (not the beta-linkage found in cellulose).

Figure \(\PageIndex{15}\): Two Primary Catalytic Mechanisms of GHs. After Payne et al. Chem. Rev. 2015, 115, 3, 1308–1448. https://doi.org/10.1021/cr500351c.
Scheme (A) shows the reaction that inverts the configuration. Water acts as a nucleophile in an SN2 reaction, with catalytic assistance by two proximal carboxylic acid side chains acting as general acids and bases. This results in an inversion of the stereochemistry at the anomeric carbon.
Scheme (B) proceeds with the retention of configuration as two different nucleophilic attacks occur. First, an active site carboxylate forms a covalent acetal intermediate with the anomeric carbon. The carboxylate, hence, acts as a nucleophilic catalyst. Water, acting as a nucleophile, then attacks to form the hemiacetal with the expulsion of the carboxylate leaving group. As discussed in Chapter 20.3 (section on glycosyl transferases), other variants of these mechanisms would include an SN1 reaction or one with an oxocarbenium-like transition state.
The CBH1 (family 7) has a long tunnel for binding cellulose. The CBH1 (TrCel7A) cellulose catalytic site spans at least nine glucose monomers (n-7, n-6,...,n-1,n+1, n+2) with cleavage typically of a cellobiose from the reducing end (between n-1 and n+1). The structure of the TrCel7A glycoside hydrolase (cellobiose hydrolase) with a small bound cellulose is shown in Figure \(\PageIndex{16}\).

Figure \(\PageIndex{16}\): Crystal structure of the first GH7 CBH and EG. Payne et al. Chem. Rev. 2015, 115, 3, 1308–1448. https://doi.org/10.1021/cr500351c Open access through Creative Commons public use license.
The ligand from the TrCel7A Michaelis complex (PDB code 4C4C (441)) is shown in all panels. (A) CBH TrCel7A CD (PDB code 1CEL (172)) view from the side, exhibiting the β sandwich structure characteristic of GH7 enzymes. TrCel7A was the first GH7 structure solved and is the best-characterized member of the GH7 family. (B) TrCel7A view from the bottom showing the more closed substrate binding "tunnel". (C) EG F. oxysporum Cel7B (PDB code 1OVW (174)) view from side. (D) FoCel7B view from the bottom showing the more open binding "groove". (E) TrCel7A Michaelis complex (PDB code 4C4C (441)) shows the standard numbering of the substrate binding sites (catalytic residues shown in green for reference). A cellulose chain enters from the −7 site. Hydrolysis occurs between the −1 and +1 sites. The +1/+2 sites are termed the "product sites".
Active site carboxylates (E212, D214, and E217) are shown near the -1/+1 cleavage site in Figure \(\PageIndex{17}\). Glu 217 is covalently attached to the -1 glucose, supporting the retaining mechanism illustrated in Fig 15 above.

Figure \(\PageIndex{17}\): Michaelis complex and glycosyl-enzyme intermediate of TrCel7A. Payne et al. ibid.
Panel (A) shows the TrCel7A Michaelis complex (PDB code 4C4C (441)).
Panel (B) shows a TrCel7A glycosyl-enzyme intermediate (PDB code 4C4D (441)) with a covalent bond between the nucleophile and the broken cellooligomer chain. There is an approximate 30° rotation of the E217 nucleophile during glycosylation.
Figure \(\PageIndex{18}\) shows a more detailed view of the first step (glycosylation f Glu 217) for the Hypocrea jecorina GH Family 7 cellobiohydrolase Cel7A

Figure \(\PageIndex{18}\): Figure 2. Glycosylation step for Hypocrea jecorina GH Family 7 cellobiohydrolase Cel7A. Knott, Brandon C. et al. - J. Am. Chem. Soc.329 (2013) https://doi.org/10.1021/ja410291u. Open access article published under an ACS AuthorChoice License
Panel (a) shows a snapshot of the reactant's conformation from a representative AS trajectory (with the substrate in green and catalytic residues in yellow) for the glycosylation step. The proton resides on Glu217.
Panel (b) shows a representative snapshot of the transition state. The −1 glucopyranose ring now adopts a different conformation.
Panel (c) shows the product of the glycosylation reaction.
Panel (d) shows a schematic view of the overall glycosylation reaction with the collective variables identified by LM colored at the transition state. The best three-component RC identified by LM includes the forming/breaking bonds involving the anomeric carbon, the breaking bond between Glu217 and its proton, and the orientation of the nucleophile Glu212.
Figure \(\PageIndex{19}\) shows the corresponding deglycosylation (of Glu 217) step.

Figure \(\PageIndex{19}\): Figure 4. Deglycosylation step results. Knott, Brandon C. et al, ibid
Panel (a) shows a snapshot of the reactant conformation from a representative AS trajectory (with the substrate in green and catalytic residues in yellow) for the deglycosylation step. The covalent glycosyl–enzyme bond is intact, and the cellobiose product is in primed GEI mode.
Panel (b) shows a representative snapshot of the transition state. Note the distorted conformation of the −1 sugar as the nucleophilic water molecule is ripped apart.
Panel (c) shows a snapshot of the product in which the glycosyl-enzyme bond has been broken, and the catalytic residues have been regenerated.
Panel (d) shows a schematic view of the overall deglycosylation reaction with the collective variables identified by LM colored at the transition state. The best three-component RC identified by LM includes the forming/breaking bonds involving the anomeric carbon, the forming/breaking bonds involving the transferring proton, and the orientation of the C3 hydroxyl of the +1 sugar.
Binding of cellulase to cellulose fibers and lignin
Many glycoside hydrolases contain distinct carbohydrate-binding domains/modules (CBD/CBM) and catalytic domains (CD). For example, TrCel7A can be cleaved by the protease papain into a 56K domain with catalytic activity on small substrates but not large cellulose ones and a smaller 10K (C-terminal) domain that itself is glycosylated and which binds to the hydrophobic surface of cellulose crystals.
In addition, many GHs have linkers connecting the catalytic domain (CD) and the carbohydrate module (CBM), which add different functions to the enzymes. The linkers vary in size and amino acid sequence. Linkers in fungi tend to be long and N- and O-glycosylated, affecting binding/catalysis. The linkers can also be intrinsically disordered, which adds dynamic complexity to their effects.
The cellulose-binding site on cellulase has been determined by solution NMR using a synthetic 36-amino-acid protein fragment from the C-terminal domain of Trichoderma reesei Cel7A (the "carbohydrate binding module or CBM"). The amino acids involved in cellohexaose (6-mer) binding were determined by perturbation of the 2D NMR structure on binding cellohexaose. As mentioned above, cellulase also binds lignin, decreasing its catalytic efficiency towards cellulose. Results of NMR binding studies of the TrCel7A carbohydrate-binding module with cellohexaose and lignins from Japanese cedar (C-MWL) and Eucalyptus globulus (E-MWL) are shown in Figure \(\PageIndex{20}\).

Figure \(\PageIndex{20}\): Comparison of interaction property between cellohexaose and MWLs. Tokunaga, Y., Nagata, T., Suetomi, T. et al. NMR Analysis on Molecular Interaction of Lignin with Amino Acid Residues of Carbohydrate-Binding Module from Trichoderma reesei Cel7A. Sci Rep 9, 1977 (2019). https://doi.org/10.1038/s41598-018-38410-9. Creative Commons Attribution 4.0 International License. http://creativecommons.org/licenses/by/4.0/.
Panel (a) shows cellohexaose bound to the flat plane surface and cleft. A triplet tyrosine (Y5, Y31, Y32) and H4, G6, Q7, I11, L28, N29, Q34, L36 define the flat plane surface.
Panel (b) shows both MWLs bound to multiple binding sites, some of which are included in the flat plane surface and cleft, even in low titrant concentrations. These non-specific binding sites are labeled green.
Figure \(\PageIndex{21}\) shows an interactive iCn3D modelof the C-terminal cellulose-binding module of cellobiohydrolase I from Trichoderma reesei (2CBH).
.png?revision=1&size=bestfit&width=412&height=370)
Figure \(\PageIndex{21}\): C-terminal cellulose-binding module of cellobiohydrolase I from Trichoderma reesei (2CBH). (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...Yva8BVMQCGdUZ7
This synthetic carbohydrate-binding module (CBM) from the C-terminal domain of cellobiohydrolase I consists of 36 residues. NMR was used to determine the structure of the CBM. 2D NMR was used to determine the amino acids interacting with cellulose. The interacting side chains are shown as sticks underneath the molecular surface (gray). The side chains are colored by hydrophobicity, with green (most hydrophobic) and yellow (somewhat hydrophobic). The numbering system refers to the 36-amino acid synthetic peptide, not the native protein. This model clearly shows that this domain binds to the hydrophobic face of the cellulose microcrystal.
The TrCel7A CBM has glycosylated serine and threonine side chains that affect binding. The interaction of the CBM with the nonpolar cellulose surface is shown in Figure \(\PageIndex{22}\).

Figure \(\PageIndex{22}\): Glycosylated TrCel7A CBM on the hydrophobic surface of cellulose. Payne et al., ibid.
Note that the aromatic groups of the triplet tyrosines, Y5, Y31, and Y32 (not labeled), are coplanar with the cellulose surface.
As mentioned above, linkers that connect the C-terminal carbohydrate-binding module (CBM) to the catalytic domain (CD) can vary in length and sequence and are also N- and O-glycosylated. Figure \(\PageIndex{23}\) shows the interactions of the glycosylated linkers with the cellulose fibers.

Figure \(\PageIndex{23}\): Molecular snapshots of TrCel7A and TrCel6A wherein the linker binds to the cellulose surface from microsecond-long MD simulations. Payne et al., ibid.
These computational predictions of cellulose linkers enhancing CBM binding to the cellulose surface were corroborated experimentally via binding isotherm measurements. N-glycosylation and O-glycosylation are shown in blue and yellow. The glycans attached to the enzyme significantly enhance cellulase binding to cellulose fibers. Payne et al. Chem. Rev. 2015, 115, 3, 1308–1448. https://doi.org/10.1021/cr500351c Open access through Creative Commons public use license
A pictorial view of the hydrolytic cleavage of cellobiose from cellulose fibers is shown in Figure \(\PageIndex{24}\).

Figure \(\PageIndex{24}\): Complete processive cycle of a GH7 CBH. TrCel7A is shown with its CD, linker, and CBM in gray "cartoon" representation. Payne et al., ibid.
N-glycosylation and O-glycosylation are shown in blue and yellow, respectively. The cellulose surface is green, and the released cellobiose product magenta. Following the CBM and CD adsorption to the substrate and initial chain threading, TrCel7A processively cleaves cellobiose from a cellulose chain end. The "Processive Cycle" includes chain processivity, hydrolysis, and product expulsion. This processive cycle repeatedly occurs until the enzyme desorbs from the cellulose surface.
Figure \(\PageIndex{25}\) shows an interactive iCn3D modelof cellulose bound to cellobiohydrolase I from Trichoderma reesei (7CEL)
.png?revision=1&size=bestfit&width=444&height=210)
Figure \(\PageIndex{25}\): Cellulose bound to cellobiohydrolase I from Trichoderma reesei (7CEL). (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...hfJFWg6hmJAxg7
Summary
(Summary written by Claude, Anthropic)
This chapter examines the biochemical barriers to cellulosic ethanol production — the conversion of lignocellulosic biomass from agricultural waste, wood, and dedicated energy crops into fermentable glucose — and the enzyme systems nature has evolved to overcome them.
Why cellulose is so resistant. Cellulose is the most abundant biomolecule on Earth and an attractive bioethanol feedstock precisely because it is not a food crop. It is a β(1,4)-linked glucose homopolymer whose glycosidic bonds have an estimated uncatalyzed hydrolysis half-life of 5 million years — far more stable than the α(1,4) bonds of starch, the phosphodiester bonds of DNA, or the amide bonds of proteins. This extraordinary stability arises from three reinforcing factors: the intrinsic chemical stability of the β-acetal bond; the crystalline packing of cellulose chains into two polymorphs (Iα and Iβ) stabilized by intralayer hydrogen bonds and interlayer hydrophobic interactions from axial C–H planes (limiting water access); and the encasing of cellulose within the secondary plant cell wall by hemicellulose (β(1,4)-backbones of xylose, mannose, or glucose, comprising 20–40% of wall mass) and lignin. About 113 gallons of ethanol can be produced per ton of corn stover — close to the 124 gallons from corn grain — making waste biomass a compelling feedstock if the enzymatic barriers can be overcome. Cellulases can increase the hydrolysis rate by up to 10¹⁷-fold over the uncatalyzed reaction without metal cofactors, making them among the most catalytically powerful enzymes known.
The lignin barrier. Lignin — up to 25% of secondary wall biomass — is synthesized from three monolignols (p-coumaryl, coniferyl, and sinapyl alcohols) derived from phenylalanine via cinnamic acid. Peroxidase enzymes generate phenoxide free radicals that couple at multiple sites to form a highly branched, heterogeneous polymer. Lignin stabilizes the cell wall, physically blocks cellulase from accessing cellulose, and also nonspecifically adsorbs cellulase through hydrophobic interactions, reducing its effective concentration available for productive hydrolysis. Thermochemical pretreatment (e.g., THF–H₂O with 0.5% H₂SO₄ at 150°C) partially erodes the biomass surface, enabling subsequent enzymatic delignification. Bacteria such as Pandoraea sp. B-6 and white-rot fungi produce two key delignifying enzymes: laccase and manganese peroxidase. Fungal laccase from Trametes versicolor (TvL) contains four copper ions organized as a T1 Cu site (near the surface in domain 3, where substrates bind) and a trinuclear T2/T3 cluster (buried at the domain 1/3 interface, where O₂ is reduced to water). Four single-electron transfers — mediated by His-458, Asp-206, Asn-264, and Phe-265 as key substrate-interaction residues — generate lignin-derived aromatic free radicals that drive ether bond cleavage, ring opening, and demethoxylation (delignification). The active site is dynamic, as confirmed by molecular dynamics simulations, accommodating a wide variety of aromatic substrates.
Cellulase mechanism and structure. Cellobiohydrolases (CBHs) are the most important cellulases industrially; Trichoderma reesei Cel7A (TrCel7A, GH family 7) is the best-characterized. It processively cleaves cellobiose (a glucose dimer) from the reducing end of a cellulose chain as it threads through a ~50 Å tunnel formed by two surface loops. The reaction proceeds by retention of anomeric configuration through a double-displacement mechanism: Glu217 acts as a nucleophile to form a covalent glycosyl-enzyme intermediate, then water attacks with general acid/base assistance from Glu212 and Asp214 to regenerate the enzyme. This contrasts with inverting glycosidases, which proceed via a single SN2-like water attack. The catalytic cycle involves initial distortion of the −1 glucopyranose ring at the transition state, followed by expulsion of cellobiose from the +1/+2 product sites.
Domain architecture and cellulose binding. TrCel7A has a modular structure: a large catalytic domain (CD) connected via a long, N- and O-glycosylated linker to a small C-terminal carbohydrate-binding module (CBM). The 36-residue CBM binds specifically to the hydrophobic face of crystalline cellulose through a flat surface defined by a triplet of tyrosine residues (Y5, Y31, Y32) that stack coplanarly against the cellulose surface via π-stacking and van der Waals interactions, supported by additional contacts from H4, G6, L28, N29, Q34, and L36. Glycosylation of the linker — both N- and O-linked sugars — significantly enhances CBM binding to cellulose fibers, as confirmed by molecular dynamics simulations and binding isotherms. The processive catalytic cycle consists of CBM and CD adsorption to the cellulose surface, initial chain threading, repeated hydrolysis and product (cellobiose) expulsion, and eventual desorption. A critical complication is that the same CBM surface that binds cellulose also binds lignin nonproductively, reducing catalytic efficiency — a key target for protein engineering to improve cellulosic ethanol yields.