
7: Algae - an Introduction
- Page ID
- 34462
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Search Fundamentals of Biochemistry
(Learning goals written by Claude, Anthropic)
By the end of this chapter, students should be able to:
Classification, Diversity, and Evolutionary Origins
- Place the major algal groups within the three-domain, seven-kingdom framework of life, explain how primary endosymbiosis (cyanobacterium uptake → green and red algae) and secondary endosymbiosis (red alga uptake → brown algae, diatoms, dinoflagellates) account for the taxonomic diversity of algae, and identify cyanobacteria as the only prokaryotic algae.
- Categorize microalgae into four practical groups (Cyanophyta, Pyrrophyta, Chrysophyta, Chlorophyta), identify representative organisms and their characteristic pigments and energy storage compounds from the classification table, and predict which groups are most suitable for bioethanol versus biodiesel production.
- Describe the structural analogs in kelp (holdfast, stipe, gas bladder, blade) compared to roots, stems, and leaves, explain why macroalgae are not plants, and identify the advantages of macroalgae over terrestrial feedstocks for biofuel production — including rapid growth (up to 30× faster than land plants), absence of lignin, and no requirement for fertilizers or agricultural land.
Light, Pigments, and Algal Depth Distribution
- Apply the relationship E = hc/λ to explain why red light is absorbed preferentially in surface waters while blue light penetrates deeper, connect this to the depth stratification of green (surface), brown (intermediate), and red (deep) algae based on their respective light-harvesting pigments, and explain the color of open ocean versus coastal waters in terms of light scattering and absorption by microorganisms.
Algal Blooms and Environmental Health
- Describe the biochemical mechanisms of toxicity of microcystin (irreversible Ser/Thr protein phosphatase inhibitor from cyanobacterial blooms) and brevetoxin (voltage-gated Na⁺ channel activator from Karenia brevis red tides), connect increasing bloom frequency to agricultural nutrient runoff and warming coastal waters, and explain the ecological consequences of bloom decay including hypoxic zone formation.
Biofuel Relevance of Major Algal Groups
- Compare the energy storage compounds of red (floridean starch, α(1,4) with α(1,6) branches), green (amylopectin-like starch in plastids), and brown (mannitol, chrysolaminarin) macroalgae, identify which are fermentable to ethanol and which require unique enzymatic strategies beyond standard amylases, and explain why the near-total absence of lignin in macroalgal cell walls makes them superior third-generation bioethanol feedstocks.
The previous chapter discussed bioethanol production from plant starches (first generation) and lignocellulosic biomass (second generation), including waste streams such as stover and sugar cane. Each had its challenges. Now, we will consider third-generation bioethanol production from algae, which has significant potential to minimize environmental damage. Below in Figure \(\PageIndex{1}\) is a summary of bioethanol production from each generation of feedstock.

Figure \(\PageIndex{x}\): Figure 1. General flowchart of bioethanol production, comparing the pre-fermentation processing of feedstocks for the first three generations of bioethanol production. The blue-highlighted area illustrates a value-added process that can enhance the value of bioethanol production. Tse, T.J.; Wiens, D.J.; Reaney, M.J.T. Production of Bioethanol—A Review of Factors Affecting Ethanol Yield. Fermentation 2021, 7, 268. https://doi.org/10.3390/fermentation7040268. Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/)
There are so many types of algae that it can be daunting to read about them. Some are single cells, some can form filaments and colonies, and some are multicellular with different cell types. Some are prokaryotes; some are eukaryotes.
The Algae Database suggests they are best defined as "oxygenic photosynthesizers other than embryophyte land plants." Naming and classification are generally based on botanical names ending with phyla = -phyta, classes = -phyceae, order = -ales, and family = -aceae. The main groups are blue-green algae (cyanobacteria), rhodophytes, phaeophytes, chlorophytes, euglenophytes, charophytes, diatoms, dinoflagellates, and cryptophytes. It's even more complicated, since some animal species have been classified as algae and have zoological name endings (-zoa, -ea, -ida, -idae). We apologize in advance for any errors or inconsistencies in the description of algae in the chapter section and ask that you contact us with any corrections.
Before we discuss biofuel production from algae, we will review the different types of algae (as we did for zooplankton and phytoplankton). Algae can be broadly divided into microalgae (seen with a microscope) and macroalgae (seen with the eye). The Algaebase lists almost 100,000 species of algae.
Here is a summary of their properties. They can
- fix carbon and produce food by photosynthesis. As such, they are primary producers.
- be tiny (microalgae) or large and visible (macroalgae, also known as seaweed). Kelp, which can form large underwater forests, is a macroalgae.
- be unicellular, form colonies or filaments, or larger multicellular structures
- attach to objects or float freely.
Some algae scientists (phytologists) consider any organism with chlorophyll but without stems, roots, leaves, flowers, or vessels to be an alga.
Algal naming and classification can be confusing, so let's start with a broad overview.
Life can be divided into domains, kingdoms, phyla, and additional subcategories. In 1990, Carl Woese proposed three domains, Archaea, Bacteria, and Eukarya, based on analyses of ribosomal RNA sequences. These domains are further classified into kingdoms: Archaea, (Eu)bacteria, Protista, Fungi, Plantae, and Animalia. A seventh kingdom was added in 1981 by Thomas Cavalier-Smith, who divided Protista into two kingdoms: Protista (unicellular eukaryotes, including some protozoa and some molds) and a new kingdom, Chromista (unicellular or multicellular eukaryotes, including algae, diatoms, and some protozoans). Both Protista and Chromista have organisms with chlorophyll, and both also have heterotrophic organisms. Newer classifications based on additional biochemical data may yet be proposed. Table \(\PageIndex{1}\) below summarizes life's domains and kingdoms.
| Domains and Kingdoms | |||||||
| Domain | Bacteria | Archaea | Eukarya | ||||
| Kingdom | (Eu)bacteria | Archaebacteria | Plantae | Animale | Fungi | Protista | Chromista |
Table \(\PageIndex{1}\): Domains and kingdoms of life.
All life arose from the last universal common ancestor, LUCA, as shown below in Figure \(\PageIndex{2}\).

Figure \(\PageIndex{2}\): Phylogenetic tree linking all major groups of living organisms to the LUCA. https://upload.wikimedia.org/wikiped..._1990_LUCA.svg
Their biological classification of algae (which illustrates how widely spread they are among different domains, kingdoms, and phyla) is shown below in Figure \(\PageIndex{3}\)

Figure \(\PageIndex{3}\): Distribution of algae among groups in the Tree of Life as recognized by the ITIS and Species 2000 (and ife.org) in 2011. The deep classification of algae is the subject of great debate, and even the higher clades have been discussed and revised recently. Adapted from Verdelho Vieira, V.; Cadoret, J.-P.; Acien, F.G.; Benemann, J. Clarification of Most Relevant Concepts Related to the Microalgae Production Sector. Processes 2022, 10, 175. https://doi.org/10.3390/pr10010175. Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Microalgae
Microalgae are single-cell organisms that can form filaments and colonies. This group has one prokaryotic member, cyanobacteria, also known as blue-green algae. As a prokaryote, it lacks mitochondria and chloroplasts. The rest of the microalgae are eukaryotic and include the phyla Chlorophyta, Rhodophyta, Glaucophyta, Cryptophyta, Euglenozoa, Cercozoa, Heterokontophyta, Haptophyta, and Miozoa (Myzozoa). Another, more straightforward organizational system divides microalgae into four categories:
- cyanophyta (blue-green algae/cyanobacteria)
- pyrrophyta (dinoflagellates and cryptomonads, and can be yellowish-green to golden-brown)
- chrysophyta (diatoms, heterokonts, and golden brown algae)
- chlorophyta (microscopic green algae). This term also applies to macroalgae (see below).
Green algae are diverse and are now divided into two phyla, Chlorophyta and Charophyta, with a combined 17 classes. AlgaeBase dynamic species counts show about 4,500 Chlorophyta species, including those living on land and in freshwater, as well as those considered macroalgal seaweeds. There are about 2500 species of Charophyta that are entirely freshwater.
Green, brown, and red algae are found within the microalgae. (There are also green, brown, and red macroscopic algae.) Microalgae are much more efficient at photosynthesis than land plants. Diatoms are a type of microalgae and are precursors to the "fossil" fuel oil deposits. Spirogyra is a unicellular green alga that forms long filaments (colonies) up to 0.1 mm in length, giving it a multicellular appearance.
One class of microalgae, the green algae, arose when a microbe acquired a cyanobacterium, enabling photosynthesis. Green algae eventually evolved into higher plants, and a similar process led to the evolution of red algae. Brown algae, diatoms, dinoflagellates, and euglenoids are other types of algae that arose from the incorporation of red or green algal lineages into other eukaryotic host cells.
Green microalgae, which absorb red wavelengths, are generally found on the surface. The red microalgae, which absorb green and blue wavelengths, are at lower levels. The brown microalgae are usually found between these water layers. Figure \(\PageIndex{4}\) below shows the light penetration spectrum in water.
The energy of light photons is given by E=hν=hc/λ, where ν is the frequency of the light, and λ is the wavelength. As highly energetic X-rays penetrate matter, visible light can penetrate water. Water absorbs incident radiation, with lower-energy, longer-wavelength photons of red light absorbed more readily in the top layers, while blue light penetrates farther into the water. The depths of penetration of light in the open ocean (left) and coastal (right) waters are shown in Figure \(\PageIndex{4}\).

Figure \(\PageIndex{4}\): The light penetration spectrum in water as a function of color. https://oceanexplorer.noaa.gov/edu/m...fact-sheet.pdf
When you are underwater, everything seems blue because red light is preferentially absorbed, leaving photons enriched in blue light to reach our eyes. Blue-enriched light reaches our eyes as it reflects off objects. Clear water in the open ocean also appears blue because there are fewer particles, such as phytoplankton, that scatter light back to our eyes. Rayleigh (or elastic light scattering) depends on 1/λ6, so low-wavelength light scatters most from particles. Sunsets and sunrises appear red when light passes through more of the atmosphere, and the blue light is scattered from atmospheric particles before reaching our eyes. Coastal waters contain more sediment, algae, and microscopic organisms, such as plankton, that can scatter light. These waters appear greener and bluer because microorganisms absorb red light, leaving blue light more scattered.
Microalgae exist as single cells or can form multicellular filaments and colonies. They proliferate in the presence of simple nutrients and can produce large amounts of polysaccharides for industrial bioethanol production or fatty acids/triacylglycerols for biodiesel production. As mentioned above, they are the source of underground oil deposits. Both freshwater microalgae (such as Chlorella and Haematococcus) and ocean microalgae (such as Dunaliella, Phaeodactylum, and Tetraselmis) can be used to produce biofuels. The cyanobacterium Spirulina sp. is commonly used commercially. It and Synechococcus have large amounts of glycogen that could be used for bioethanol production. Their triacylglycerol content is low, so they can't be used for biodiesel (which we will discuss in a later section of the chapter).
Most are familiar with algae blooms in freshwater lakes (and even in saltwater environments). Cyanobacteria (blue-green algae) are the main culprit. Microcystin, a potentially lethal toxin that targets serine/threonine protein phosphatases, is released from some algal blooms. Figure \(\PageIndex{5}\) shows a microalgae bloom in Lake Erie, a shallow freshwater lake polluted with agricultural runoff, making it an excellent site for cyanobacterial blooms.

Figure \(\PageIndex{5}\): Microalgae bloom in Lake Erie, October 2011. https://commons.wikimedia.org/wiki/F..._Lake_Erie.jpg
Red tides, an informal name for harmful algal blooms, occur in coastal waters. Along the US coast, they are caused by dinoflagellates, diatoms, and cyanobacteria. Around the Gulf of Mexico, the leading cause of red tides is the microalgae Karenia brevis. They release large amounts of brevotoxin, a polycyclic ether that binds to and activates voltage-gated Na+ channels in nerve and muscle. Hence, it is a potentially deadly neurotoxin. The frequency of red tides is increasing worldwide. Figure \(\PageIndex{6}\) shows two images of red tides.
 |
 |
Figure \(\PageIndex{6}\): Red Tide. License from Shutterstock
Microalgae are sources of glycan polymers that can be used for bioethanol production; some are excellent candidates for biodiesel fuel made from triacylglycerol reserves. Table \(\PageIndex{2}\) below shows a classification of microalgae and Cyanobacteria along with their characteristic pigments (that impart their distinctive colors) and their energy reserves that could be used for biofuel production.
| Phylum | Class | Pigments | Reserve | Habitat |
|---|---|---|---|---|
| Cyanobacteria | Cyanophyceae | Chl a, β-carotene, flavacene, Echinenone isozea-, zea-, myxo-, oscillaxanthin APC, C-PC, C-PE |
Starch (granule) and glycogen |
Marine Freshwater Terrestrial |
| Euglenophyta | Euglenophyceae | Some colorless Chl a, b, diadinoxanthin |
Paramylon Ergosterol |
Marine Freshwater Terrestrial |
| Heterokontophyta/ Ochrophyta |
Xanthophyceae Eustigmatophyceae |
Chl a and c, β-carotene, heteroxanthin, diadinoxanthin (++) | Oil Leucosin Ergosterol |
Marine Freshwater Terrestrial |
| Miozoa | Dinophyceae | Chl a, c, β-carotene, diadinoxanthin, dinoxanthin, peridinins | Starch Lipids |
Marine Freshwater |
| Heterokontophyta/ Ochrophyta |
Chrysophycea | Chl a, c, β-carotene, Fuco-, Diato-, diadinoxanthin |
Chrysolaminarin Fucosterol Porifasterol |
Marine Freshwater |
| Haptophyta | Coccolithophyceae Pavlovophyceae Rappephyceae |
Chl a, c, β-carotdnd, Fuco-, Diato-, diadinoxanthin |
Chrysolaminarin Fucosterol Porifasterol |
Marine Freshwater |
| Bacillariophyta (Diatoms) | Bacillariophyceae | Chl a, c, β-carotdnd, Fuco-, Diato-, diadinoxanthin |
Chrysolaminarin Oil |
Marine Freshwater Terrestrial |
| Cryptophyta | Cryptophyceae | Chl a, c, Biliproteins, α-carotene, Allo-, Croco-, Monado-xanthin | Starch (granule) Oil Carbohydrates |
Marine Freshwater |
Table \(\PageIndex{2}\): Classification of microalgae and Cyanobacteria. Hachicha, R.; Elleuch, F.; Ben Hlima, H.; Dubessay, P.; de Baynast, H.; Delattre, C.; Pierre, G.; Hachicha, R.; Abdelkafi, S.; Michaud, P.; Fendri, I. Biomolecules from Microalgae and Cyanobacteria: Applications and Market Survey. Appl. Sci. 2022, 12, 1924. https://doi.org/10.3390/app12041924. Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Cyanobacteria and red microalgae store glycogen and floridean starch (a hybrid between starch and glycogen), respectively, while green microalgae accumulate amylopectin-like polysaccharides.
Macroalgae
As mentioned above, macroalgae, known as seaweeds, lack roots, stems, leaves, or flowers. Kelp in underwater forests appears to have roots, stems, and leaves, but it is not a plant. They have analogous structures, such as holdfasts, stipes, and blades, which serve the functions of roots, stems, and leaves found in plants. As with microalgae, variants of macroalgae include red (Rhodophyta), green (Chlorophyta, also a type of microalgae), and brown (Phaeophyta) algae. They grow up to 30 times as quickly as land-based groups. In addition, they lack lignin. They grow much faster than terrestrial plants and can produce large amounts of biomass for commercial processing on much less land than land-based crops. They can be grown cheaply in sea farms without adding nutrients or pesticides. Hence, they are ideal for both food and biofuel production. Brown and red algae have many present commercial uses. They have abundant carbohydrates for potential bioethanol production and triacylglycerols for biodiesel production. Their carbohydrate composition includes mannitol and cell wall constituents, which could also be used for fermentation. In contrast to present petroleum sources, the biodiesel from macroalgae does not contain sulfur.
Red macroalgae
(some material below for red and green macroalgae from https://bio.libretexts.org/Bookshelv...nd_Green_Algae)
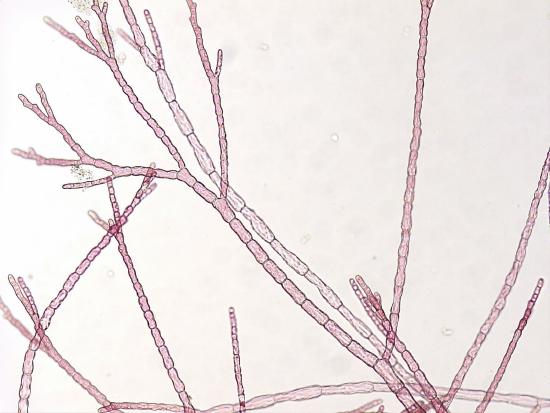
The red algae are almost exclusively marine; some are unicellular, but most are multicellular. They have true chloroplasts with two membranes (no residual peptidoglycan) that contain chlorophyll. Like cyanobacteria, they use phycobilins as antenna pigments: phycoerythrin (which gives them a red color) and phycocyanin. Red pigment allows red algae to photosynthesize at greater depths than green or brown algae, harnessing more of the blue light that penetrates deeper into the water column. Unlike green algae and plants, red algae store carbohydrates as Floridean starch, which has glucose in α(1,4) linkages and occasional α(1,6) linkages, similar to amylopectin. Agar, the base for culturing bacteria and other microorganisms, is extracted from a red alga. Multicellular forms can be filamentous, leafy, sheet-like, coralloid, or crust-like. Some examples are shown in Figure \(\PageIndex{7}\).
 |
 |
 |
Figure \(\PageIndex{7}\): These images show multicellular red algae, which can range from filamentous (first image) to "leafy" (second image, left) to sheet-like (second image, right). The red color is due to an abundance of the red pigment phycoerythrin, which gives this group reddish chloroplasts. First image by Melissa Ha CC-BY-NC. Second image by Maria Morrow CC-BY-NC. Right image: https://commons.wikimedia.org/wiki/F...ed_algae_3.jpg
Green macroalgae
These algae exhibit a great diversity of form and function. Similar to red algae, green algae can be unicellular or multicellular. Many unicellular species form colonies, and some green algae exist as large, multinucleated single cells. Green algae primarily inhabit freshwater and damp soil and are a common component of plankton. They have chloroplasts and the photosynthetic pigments chlorophyll a and b, carotene, and xanthophylls. Examples include Chlamydomonas, Chlorella, Pediastrum, Netrium, Hydrodictyon, Acetabularia, Ulva, and Spirogyra. Lichens are a symbiotic combination of fungi and green algae.
The nature of evolutionary relationships among green algae remains debated. As of 2019, genetic data support splitting the green algae into two major lineages: chlorophytes and streptophytes. The green algae exhibit features similar to those of land plants, particularly in chloroplast structure. They have chlorophyll a and b, have lost phycobilins but gained carotenoids, and store carbohydrates as starch inside plastids. Green algae are an important food source for many aquatic animals. Figure \(\PageIndex{8}\) shows two types of green macroalgae.
 |
 |
Figure \(\PageIndex{8}\):Two types of green macroalgae
Left: Figure 5.3.3.125.3.3.12: Trentepohlia is a genus of green algae in terrestrial environments. It forms fluffy orange colonies on trees and is a photobiont in many lichens. One might not realize they were looking at green algae because of the orange pigmentation. However, green algae have carotenoids. These terrestrial green algae produce abundant carotenoids, perhaps for protection against sun damage. Photo by Scott Loarie, CC0.
Right: freshwater green algae. https://upload.wikimedia.org/wikiped...reen_Algae.jpg
Green macroalgal blooms (called green tides) can also occur (just as blooms from the microalgae cyanobacteria). Green tides in the Yellow Sea (between China and Korea) are the largest known. A particularly large one is shown in Figure \(\PageIndex{9}\):

Figure \(\PageIndex{9}\): Green Seaweed in the Yellow Sea , June 2021. https://earthobservatory.nasa.gov/im...the-yellow-sea
This particular green color was from the nontoxic green macroalgae Ulva prolifera. Because it is edible, it is often called sea lettuce. Blooms can affect the local ecosystem and, as they decay, lead to hypoxic zones.
Brown macroalgae (also known as Kelp)
(Some material below for brown macroalgae from https://bio.libretexts.org/Bookshelv...3A_Brown_Algae)
Macroscopic brown algae arose when a heterotrophic eukaryote merged with a unicellular photosynthetic eukaryotic red alga's chloroplasts. This time, the red alga became a chloroplast with four membranes -- the engulfing membrane from the oomycete, the red alga's plasma membrane, and the two membranes of the original chloroplast within the red alga. The chloroplast has lost one of these membranes in many groups derived from secondary endosymbiosis. Figure \(\PageIndex{10}\) shows this secondary endosymbiosis event.
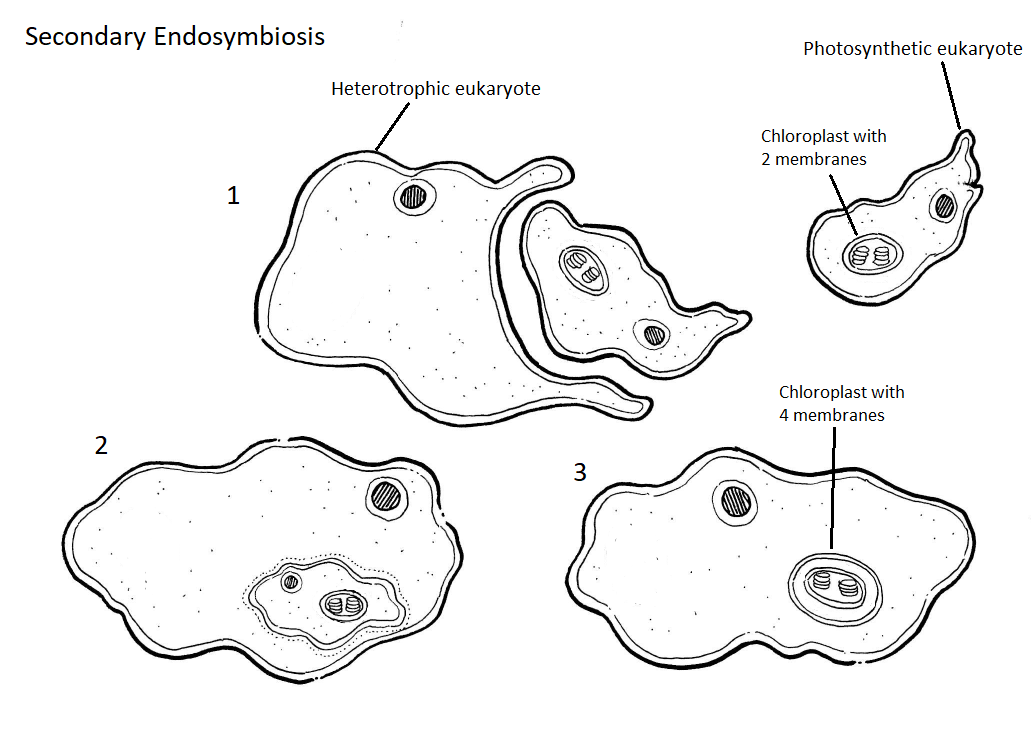
Figure \(\PageIndex{10}\): In the diagram above, we see a unicellular photosynthetic eukaryote with a 2-membrane chloroplast. In step one, this organism is engulfed by a heterotrophic eukaryote. In step two, we see the photosynthetic organism inside the heterotrophic organism. In step three, the original photosynthetic organism has been reduced to a chloroplast with four membranes. Artwork by Nikki Harris CC-BY-NC with added labels by Maria Morrow.
Brown algae are brown because they produce large amounts of carotenoids, primarily fucoxanthin. These organisms are exclusively multicellular and can grow so large that they require specialized conductive cells to transport the products of photosynthesis from their blades to the rest of their tissues. These conductive cells, called trumpet hyphae, have sieve plates and resemble sieve tubes found in flowering plants.
Much like Saprolegnia, the body of an alga is called a thallus because it is not differentiated into specialized tissues. The general morphology of a brown alga includes a holdfast, stipe, gas bladder(s), and blade(s). Figure \(\PageIndex{111}\) shows a diagram of the kelp structure.
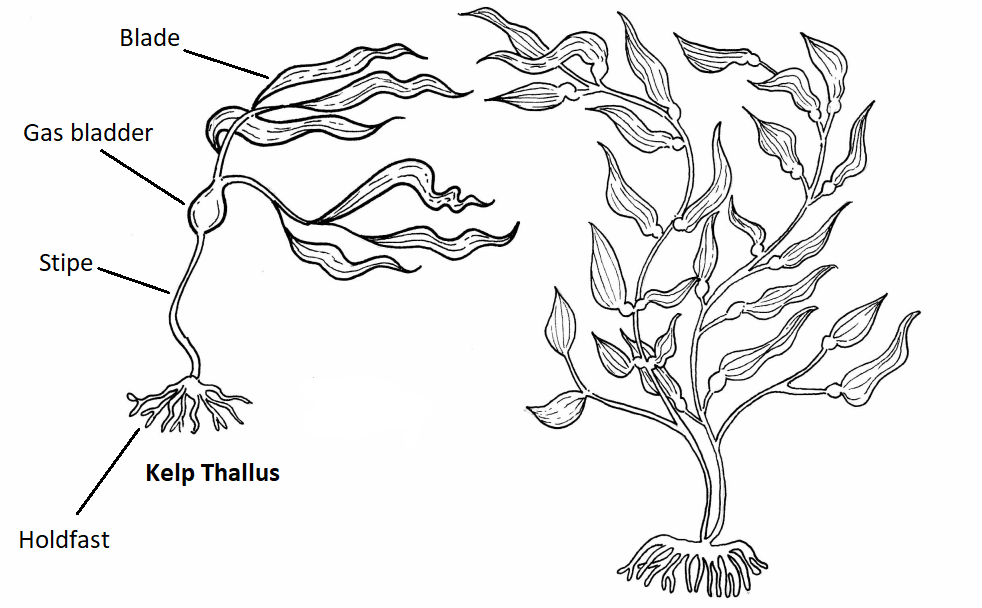
Figure \(\PageIndex{11}\): A diagram of kelp structure. In the diagram above, there are two kelp thalli. The one on the left side is labeled. At the base of the thallus is a network of root-like projections that form the holdfast. The stem-like structure that travels up from the holdfast is the stipe, which terminates in an inflated gas bladder. There are several leaf-like structures attached to the gas bladder. These are blades. The thallus on the right has all these components but in a slightly different arrangement. Can you find them? Artwork by Nikki Harris CC-BY-NC with added labels by Maria Morrow.
Figure \(\PageIndex{12}\) shows a beautiful image of a kelp forest.
.jpg?revision=1)
Figure \(\PageIndex{12}\): Kelp forest. https://commons.wikimedia.org/wiki/F...1115735%29.jpg
Figure \(\PageIndex{13}\) shows another type of brown macroalga.
.jpg?revision=1&size=bestfit&width=349&height=262)
Figure \(\PageIndex{12}\): Underwater "roses" of a brown algae Padina pavonica, commonly known as the peacock's tail (Israel). https://commons.wikimedia.org/wiki/F...l_(Israel).jpg
Now we can discuss algae as a source of biofuels and nutritious foods in the next chapter section.
Summary
(Summary written by Claude, Anthropic)
This chapter introduces the remarkable diversity of algae — the primary photosynthetic organisms of aquatic environments — as a foundation for understanding their potential as third-generation biofuel feedstocks and as key players in the global carbon cycle.
Classification and evolutionary origins. Algae are best defined as oxygenic photosynthesizers other than embryophyte land plants — a functional grouping that spans multiple domains, kingdoms, and phyla. The ~100,000 known species are distributed across the bacterial domain (cyanobacteria, the only prokaryotic algae) and multiple eukaryotic kingdoms (Protista, Chromista, and Plantae). Their evolutionary diversity reflects repeated endosymbiotic events: green and red algae arose through primary endosymbiosis when a heterotrophic eukaryote engulfed a cyanobacterium, retaining it as a two-membrane chloroplast; brown algae, diatoms, and dinoflagellates arose through secondary endosymbiosis when a heterotrophic eukaryote engulfed a photosynthetic red alga, producing a chloroplast with three or four membranes. Green algae eventually gave rise to all land plants. Microalgae are conveniently grouped into four categories: cyanophyta (blue-green/cyanobacteria), pyrrophyta (dinoflagellates, cryptomonads), chrysophyta (diatoms, golden brown algae), and chlorophyta (green microalgae). The AlgaeBase lists approximately 100,000 algal species overall.
Light penetration and algal depth distribution. Because photon energy is inversely proportional to wavelength (E = hc/λ), shorter-wavelength blue light carries more energy and penetrates more deeply in water than longer-wavelength red light, which is preferentially absorbed in the upper water column. Green algae, which absorb red wavelengths via chlorophyll a and b, are most productive near the surface. Red algae, which absorb green and blue wavelengths using phycoerythrin and phycocyanin as accessory pigments, photosynthesize efficiently at greater depths. Brown algae occupy intermediate depths, using fucoxanthin to capture the blue-green light available there. This stratification explains both the color of ocean water and the zonation of algal communities in marine environments.
Microalgae. Microalgae are unicellular or colonial organisms distinguished by rapid growth, high photosynthetic efficiency, and diverse energy storage compounds. Cyanobacteria and red microalgae store glycogen and floridean starch (α(1,4) with α(1,6) branches, similar to amylopectin) respectively; green microalgae accumulate amylopectin-like polysaccharides. Diatoms and some heterokont algae store chrysolaminarin and oils; dinoflagellates store starch and lipids. Species such as Chlorella, Spirulina, Dunaliella, and Tetraselmis are candidates for bioethanol and biodiesel production from their carbohydrate and triacylglycerol reserves respectively. Diatoms are the ancestral source of the petroleum deposits that now fuel the fossil fuel economy. Microalgal blooms are an increasing environmental concern: cyanobacterial blooms in nutrient-polluted freshwater (such as Lake Erie) release microcystin, a cyclic peptide toxin that irreversibly inhibits serine/threonine protein phosphatases; coastal red tides (caused by Karenia brevis in the Gulf of Mexico) release brevetoxin, a polycyclic ether that activates voltage-gated Na⁺ channels in nerve and muscle, causing potentially fatal paralytic shellfish poisoning. The increasing frequency of both bloom types is linked to agricultural nitrogen runoff and climate-driven warming of coastal waters.
Macroalgae. Macroalgae (seaweeds) are multicellular, visible to the naked eye, and lack the roots, stems, leaves, and vascular systems of true plants. Their structural analogs — holdfasts, stipes, gas bladders, and blades — serve equivalent mechanical and physiological functions. Red macroalgae (Rhodophyta) are almost exclusively marine, store floridean starch, and use phycoerythrin for deep-water photosynthesis; agar, derived from red algae, is essential for microbial culture media. Green macroalgae (Chlorophyta) inhabit primarily freshwater and coastal zones, store starch in plastids, and include ecologically important species such as Ulva (sea lettuce). Brown macroalgae (Phaeophyta/kelp) are exclusively multicellular, can form towering underwater forests, and are colored by fucoxanthin; their largest forms contain specialized conductive cells (trumpet hyphae) analogous to plant sieve tubes for long-distance transport of photosynthate.
Macroalgae offer compelling advantages as biofuel feedstocks: they grow up to 30 times faster than terrestrial plants, contain no lignin (eliminating the most significant barrier to cellulosic ethanol), require no agricultural land, and need no added nutrients or pesticides when farmed in the ocean. Their carbohydrates — including mannitol, laminarin, floridean starch, and cell wall polysaccharides — are fermentable to ethanol, and their triacylglycerol content can be processed to biodiesel without the sulfur contamination inherent in petroleum-derived fuels. These properties position macroalgae as a highly promising third-generation biofuel feedstock, setting the stage for the detailed treatment of algal bioethanol and biodiesel production in subsequent sections of the chapter.