
8.7: Mesozoic
- Page ID
- 8562


Following the Permian Mass Extinction, the Mesozoic (“middle life”) was from 252 million years ago to 66 million years ago. As Pangea started to break apart, mammals, birds, and flowering plants developed. The Mesozoic is probably best known as the age of reptiles, most notably, the dinosaurs.
Mesozoic Tectonics and Paleogeography

Pangea started breaking up (in a region that would become eastern Canada and United States) around 210 million years ago in the Late Triassic. Clear evidence for this includes the age of the sediments in the Newark Supergroup rift basins and the Palisades sill of the eastern part of North America and the age of the Atlantic ocean floor. Due to sea-floor spreading (Chapter 3), the oldest rocks on the Atlantic’s floor are along the coast of northern Africa and the east coast of North America, while the youngest are along the mid-ocean ridge.
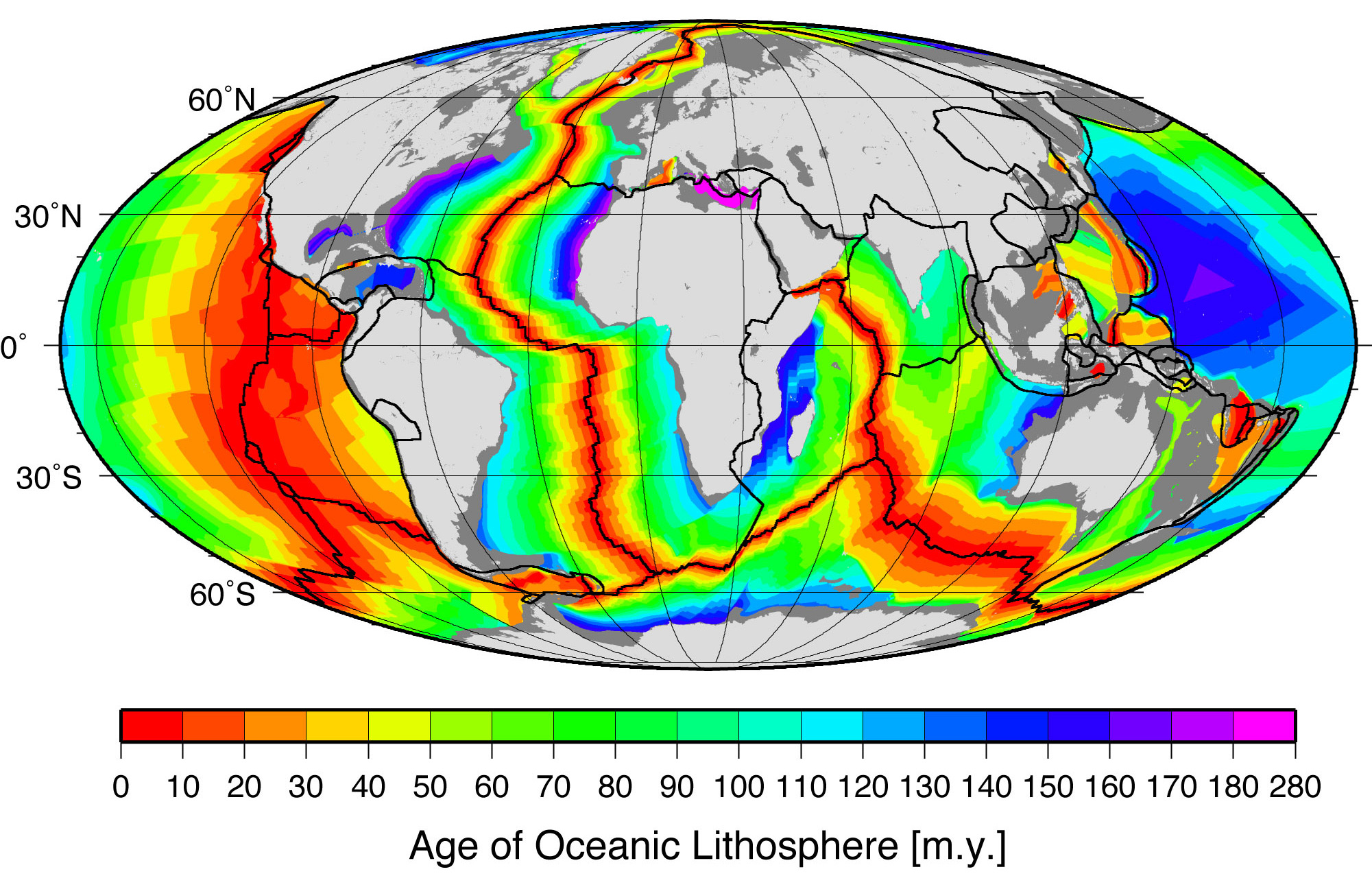
This age pattern shows how the Atlantic Ocean opened as the young Mid-Atlantic Ridge began to create the seafloor. This means the Atlantic ocean started opening and was first formed here. The southern Atlantic opened next, with South America separating from central and southern Africa. Last (happening after the Mesozoic ended) was the northernmost Atlantic, with Greenland and Scandinavia parting ways. The breaking points of each rifted plate margin eventually turned into the passive plate boundaries of the east coast of the Americas today
Video of Pangea breaking apart and plates moving to their present locations. By Tanya Atwater.
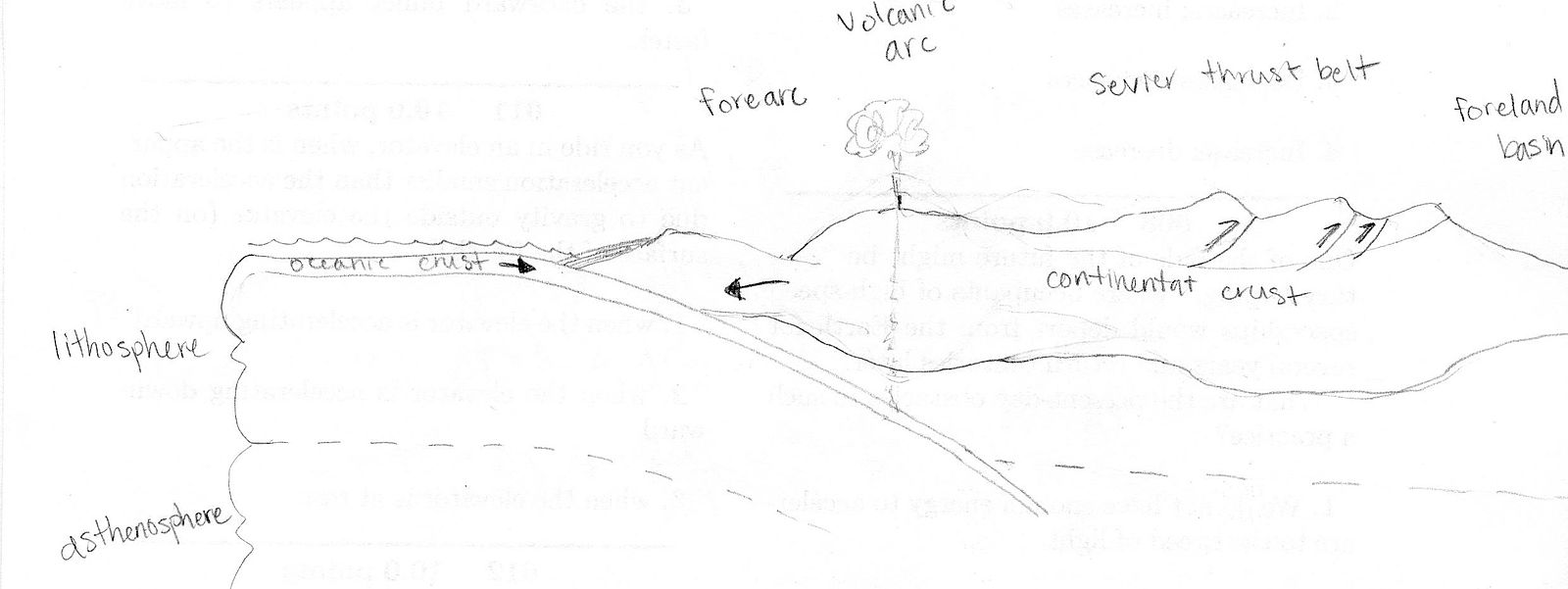
In western North America, an active plate margin had started with subduction, controlling most of the tectonics of that region in the Mesozoic. Another possible island-arc collision created the Sonoman Orogeny in Nevada during the latest Paleozoic to the Triassic. In the Jurassic, another island-arc collision caused the Nevadan Orogeny, a large Andean-style volcanic arc and thrust belt [105]. The Sevier Orogeny followed in the Cretaceous, which was mainly a volcanic arc to the west and a thin-skinned fold and thrust belt to the east, meaning stacks of shallow faults and folds built up the topography. Many of the structures in the Rocky Mountains today date from this orogeny.
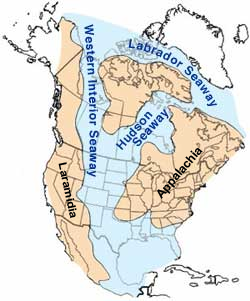
Tectonics had an influence in one more important geographic feature in North America: the Cretaceous Western Interior Foreland Basin, which flooded during high sea levels forming the Cretaceous Interior Seaway. Subduction from the west was the Farallon Plate, an oceanic plate connected to the Pacific Plate (seen today as remnants such as the Juan de Fuca Plate, off the coast of the Pacific Northwest). Subduction was shallow at this time because a very young, hot and less dense portion of the Farallon plate was subducted. This shallow subduction caused a down warping in the central part of North America [107]. High sea levels due to shallow subduction, and increasing rates of seafloor spreading and subduction, high temperatures, and melted ice also contributed to the high sea levels [108]. These factors allowed a shallow epicontinental seaway that extended from the Gulf of Mexico to the Arctic Ocean to divide North America into two separate landmasses [109], Laramidia to the west and Appalachia to the east, for 25 million years [59]. Many of the coal deposits in Utah and Wyoming formed from swamps along the shores of this seaway [111]. By the end of the Cretaceous, cooling temperatures caused the seaway to regress [112].
Mesozoic Evolution
 [<a data-cke-saved-href=\"http://creativecommons.org/licenses/by-sa/2.5\" href=\"http://creativecommons.org/licenses/by-sa/2.5\"") CC BY-SA 2.5], via Wikimedia Commons" width="372px" height="279px" src="/@api/deki/files/7956/Mesozoic_Scene-300x225.jpg">
CC BY-SA 2.5], via Wikimedia Commons" width="372px" height="279px" src="/@api/deki/files/7956/Mesozoic_Scene-300x225.jpg">The Mesozoic era is dominated by reptiles, and more specifically, the dinosaurs. The Triassic saw devastated ecosystems that took over 30 million years to fully re-emerge after the Permian Mass Extinction [113]. The first appearance of many modern groups of animals that would later flourish occurred at this time. This includes frogs (amphibians), turtles (reptiles), marine ichthyosaurs and plesiosaurs (marine reptiles), mammals, and the archosaurs. The archosaurs (“ruling reptiles”) include ancestral groups that went extinct at the end of the Triassic, as well as the flying pterosaurs, crocodilians, and the dinosaurs. Archosaurs, like the placental mammals after them, occupied all major environments: terrestrial (dinosaurs), in the air (pterosaurs), aquatic (crocodilians) and even fully marine habitats (marine crocodiles). The pterosaurs, the first vertebrate group to take flight, like the dinosaurs and mammals, start small in the Triassic.
 (Own work) [<a data-cke-saved-href=\"http://www.gnu.org/copyleft/fdl.html\" href=\"http://www.gnu.org/copyleft/fdl.html\"") GFDL or CC BY 3.0], via Wikimedia Commons" width="404px" height="255px" src="/@api/deki/files/7959/Augustasaurus_BW-300x189.jpg">
GFDL or CC BY 3.0], via Wikimedia Commons" width="404px" height="255px" src="/@api/deki/files/7959/Augustasaurus_BW-300x189.jpg">At the end of the Triassic, another mass extinction event occurred [114], the fourth major mass extinction in the geologic record. This was perhaps caused by the Central Atlantic Magmatic Province flood basalt [115]. The end-Triassic extinction made certain lineages go extinct and helped spur the evolution of survivors like mammals, pterosaurs (flying reptiles), ichthyosaurs/plesiosaurs/mosasaurs (marine reptiles), and dinosaurs.
 [<a data-cke-saved-href=\"http://creativecommons.org/licenses/by-sa/4.0\" href=\"http://creativecommons.org/licenses/by-sa/4.0\"") CC BY-SA 4.0], via Wikimedia Commons" width="364px" height="243px" src="/@api/deki/files/7961/Megazostrodon_sp._Natural_History_Museum_-_London-300x200.jpg">
CC BY-SA 4.0], via Wikimedia Commons" width="364px" height="243px" src="/@api/deki/files/7961/Megazostrodon_sp._Natural_History_Museum_-_London-300x200.jpg">Mammals, as previously mentioned, got their start from a reptilian synapsid ancestor possibly in the late Paleozoic. Mammals stayed small, in mainly nocturnal niches, with insects being their largest prey. The development of warm-blooded circulation and fur may have been a response to this lifestyle [118].
In the Jurassic, species that were previously common flourished due to a warmer and more tropical climate. The dinosaurs were relatively small animals in the Triassic period of the Mesozoic but became truly massive in the Jurassic. Dinosaurs are split into two groups based on their hip structure [120], i.e. orientation of the pubis and ischium bones in relationship to each other. This is referred to as the “reptile hipped” saurischians and the “bird-hipped” ornithischians. This has recently been brought into question by a new idea for dinosaur lineage [121].
Most of the dinosaurs of the Triassic were saurischians, but all of them were bipedal. The major adaptive advantage dinosaurs had was changes in the hip and ankle bones, tucking the legs under the body for improved locomotion as opposed to the semi-erect gait of crocodiles or the sprawling posture of reptiles. In the Jurassic, limbs (or a lack thereof) were also important to another group of reptiles, leading to the evolution of Eophis, the oldest snake.
 (Own work) [<a data-cke-saved-href=\"http://creativecommons.org/licenses/by-sa/3.0\" href=\"http://creativecommons.org/licenses/by-sa/3.0\"") CC BY-SA 3.0], via Wikimedia Commons" width="359px" height="189px" src="/@api/deki/files/7971/Beipiao1mmartyniuk-300x158.png">
CC BY-SA 3.0], via Wikimedia Commons" width="359px" height="189px" src="/@api/deki/files/7971/Beipiao1mmartyniuk-300x158.png">There is a paucity of dinosaur fossils from the Early and Middle Jurassic but by the Late Jurassic, they were dominating the planet. The saurischians diversified into the giant herbivorous (plant-eating) long-necked sauropods weighing up to 100 tons and bipedal carnivorous theropods, with the possible exception of the Therizinosaurs [125]. All of the ornithischians (e.g Stegosaurus, Iguanodon, Triceratops, Ankylosaurus, Pachycephalosaurus) were herbivorous with a strong tendency to have a “turtle-like” beak at the tips of their mouths.
 (Own work) [<a data-cke-saved-href=\"http://creativecommons.org/licenses/by-sa/3.0\" href=\"http://creativecommons.org/licenses/by-sa/3.0\"") CC BY-SA 3.0 or GFDL], via Wikimedia Commons" width="243px" height="328px" src="/@api/deki/files/7962/Archaeopteryx_lithographica_Berlin_specimen-222x300.jpg">
CC BY-SA 3.0 or GFDL], via Wikimedia Commons" width="243px" height="328px" src="/@api/deki/files/7962/Archaeopteryx_lithographica_Berlin_specimen-222x300.jpg">The pterosaurs grew and diversified in the Jurassic, and another notable arial organism developed and thrived in the Jurassic: birds. When Archaeopteryx was found in the Solnhofen Lagerstätte of Germany, a seeming dinosaur-bird hybrid, it started the conversation on the origin of birds. The idea that birds evolved from dinosaurs occurred very early in the history of research into evolution, only a few years after Darwin’s On the Origin of Species [127]. This study used a remarkable fossil of Archeopteryx from a transitional animal between dinosaurs and birds. Small meat-eating theropod dinosaurs were likely the branch that became birds due to their similar features [128]. A significant debate still exists over how and when powered flight evolved. Some have stated a running-start model [129], while others have favored a tree-leaping gliding model or even a semi-combination: flapping to aid in climbing.
 [<a data-cke-saved-href=\"http://www.gnu.org/licenses/old-licenses/fdl-1.2.html\" href=\"http://www.gnu.org/licenses/old-licenses/fdl-1.2.html\"") GFDL 1.2 or FAL], via Wikimedia Commons" width="219px" height="330px" src="/@api/deki/files/7975/Argentinosaurus_DSC_2943-199x300.jpg">
GFDL 1.2 or FAL], via Wikimedia Commons" width="219px" height="330px" src="/@api/deki/files/7975/Argentinosaurus_DSC_2943-199x300.jpg">The Cretaceous saw a further diversification, specialization, and domination of the dinosaurs and other fauna. One of the biggest changes on land was the transition to angiosperm-dominated flora. Angiosperms, which are plants with flowers and seeds, had originated in the Cretaceous [132], switching many plains to grasslands by the end of the Mesozoic [133]. By the end of the period, they had replaced gymnosperms (evergreen trees) and ferns as the dominant plant in the world’s forests. Haplodiploid eusocial insects (bees and ants) are descendants from Jurassic wasp-like ancestors that co-evolved with the flowering plants during this time period. The breakup of Pangea not only shaped our modern world’s geography but biodiversity at the time as well. Throughout the Mesozoic, animals on the isolated, now separated island continents (formerly parts of Pangea), took strange evolutionary turns. This includes giant titanosaurian sauropods (Argentinosaurus) and theropods (Giganotosaurus) from South America.
K-T Extinction
Similar to the end of the Paleozoic era, the Mesozoic Era ended with the K-Pg Mass Extinction (previously known as the K-T Extinction) 66 million years ago [136]. This extinction event was likely caused by a large bolide (an extraterrestrial impactor such as an asteroid, meteoroid, or comet) that collided with earth. Ninety percent of plankton species, 75% of plant species, and all the dinosaurs went extinct at this time.
 via Wikimedia Commons" width="300" src="/@api/deki/files/7967/Impact_event-300x209.jpg">
via Wikimedia Commons" width="300" src="/@api/deki/files/7967/Impact_event-300x209.jpg">One of the strongest pieces of evidence comes from the element iridium. Quite rare on Earth, and more common in meteorites, it has been found all over the world in higher concentrations at a particular layer of rock that formed at the time of the K-T boundary. Soon other scientists started to find evidence to back up the claim. Melted rock spheres [138], a special type of “shocked” quartz called stishovite, that only is found at impact sites, was found in many places around the world. The huge impact created a strong thermal pulse that could be responsible for global forest fires [141], strong acid rains [142], a corresponding abundance of ferns, the first colonizing plants after a forest fire [143], enough debris thrown into the air to significantly cool temperatures afterward [144; 145], and a 2-km high tsunami inferred from deposits found from Texas to Alabama.
 via Wikimedia Commons" width="307px" height="344px" src="/@api/deki/files/7970/Chicxulub_radar_topography-268x300.jpg">
via Wikimedia Commons" width="307px" height="344px" src="/@api/deki/files/7970/Chicxulub_radar_topography-268x300.jpg">Still, with all this evidence, one large piece remained missing: the crater where the bolide impact. It was not until 1991 that the crater was confirmed using petroleum company geophysical data. Even though it is the third-largest confirmed crater on Earth at roughly 180 km wide, the Chicxulub Crater was hard to find due to being partially underwater and partially obscured by the dense forest canopy of the Yucatan Peninsula. Coring of the center of the impact called the peak ring contained granite, indicating the impact was so powerful that it lifted basement sediment from the crust several miles toward the surface [149]. In 2010, an international team of scientists reviewed 20 years of research and blamed the impact for the extinction [150].
With all of this information, it seems like the case would be closed. However, there are other events at this time which could have partially aided the demise of so many organisms. For example, sea levels are known to be slowly decreasing at the time of the K-T event, which is tied to marine extinctions [151], though any study on gradual vs. sudden changes in the fossil record is flawed due to the incomplete nature of the fossil record [152]. Another big event at this time was the Deccan Traps flood basalt volcanism in India. At over 1.3 million cubic kilometers of material, it was certainly a large source of material hazardous to ecosystems at the time, and it has been suggested as at least partially responsible for the extinction [153]. Some have found the impact and eruptions too much of a coincidence, and have even linked the two together [154].